| Гостевая | yurishmukler@yahoo.com | Ю. Б. Шмуклер | Галерея | О себе |
|---|---|---|---|---|
| Новости | Летопись текстов | Из дальних странствий | Моя семья и другие звери | |
| Биологическое | ЦСКА | Друзья и родственники | Генеалогия | Бреды и анекдоты |
| Хохмы и афоризмы | В сущности оранжевый... | Стишки и хокку | Долгое падение на камни | Чашечка кофе |
| 1 | 2 | 3 | 4 |
Долгое падение на камниТоржественная порка Результатами экспедиции я был вдохновлен до чрезвычайности. И это – не щенячий восторг, потому что и «старшие товарищи» из ИППИ проявляли энтузиазм, и считали, что наши результаты достойны публикации – все-таки у нас получилось влиять на судьбу изолированных клеток и нашлось доказательство существования ранних межклеточных взаимодействий. Собственно говоря, так оно и оказалось впоследствии и подтвердилось множеством экспериментов, но тогда-то их у нас не было, и я и сам не представлял, куда меня все это заведет. При этом Чайлахян с самого начала дистанцировался от моей тематики с серотонином – ни тогда, ни, по-моему, потом он в трансмиттерные регуляторные эффекты не верил, а, может просто ими не интересовался. Движимый стремлением поделиться «великим открытием», я попросил ГенСеича организовать физиологический коллоквиум с приглашением лаборатории эмбриологии. Помнится, набрался почти полный конференц-зал, и, в числе прочих СанСаныч Нейфах, Анна Самойловна Гинзбург, Маша Скоблина, Васецкий, ну, и физиологи in corpore. Собственно, вся эта затея с семинаром – памятник моей самонадеянности и невежественности в основах эмбриологии. Единственное, что меня, если не извиняет, то не выставляет полным идиотом – это то, что я был в неплохой компании… Дело в том, что в принципе о классических опытах по изоляции бластомеров я, конечно, слышал, но в самом общем виде – что это возможно. Насколько я помню, раннее развитие, как и сейчас, в университетском курсе эмбриологии затрагивали только кратко и очень поверхностно – как нечто очевидное и само собой разумеющееся. Сам я никогда оригинальных трудов эмбриологических основоположников до того не читал… И никто из тех, кто работал в той экспедиции – тоже. Если бы хоть кто-нибудь был обременен классическими знаниями, мы бы эту работу даже не начинали – там, как выяснилось, уже всё всем давным-давно известно. Sciencia consomatum est![1] Вообще-то, если задуматься – с чем я вылез крушить классическую теорию, не зная, что крушу ее? У нас было по 3 – 4 десятка изолированных бластомеров на каждом из трех видов морских ежей, причем четкое распадение типов дробления надежно было обнаружено только на двух. Эффект серотонина, вообще был к тому времени на полутора десятках бластомеров только одного вида показан. Я уж и не говорю о теоретической подготовке… На пальцах, потому что, помнится, никаких слайдов еще не было, я объяснил полученные эффекты, привел свою «богатую» статистику, изложил нашу идею о периоде, в ходе которого происходит межклеточный обмен сигналами, и стал ждать похвал. Вот чего я не дождался, так это похвал. Встала Маша Скоблина и страстно объяснила дураку, что просто «когда мы получаем равномерное деление – происходит регуляция, а когда получаем микромеры – регуляции не происходит». Здесь я еще сохранял реактивность и ответил, что мы и пытаемся объяснить, в чем эта «регуляция» состоит. Что, возможно, серотониновый сигнал и представляет собой именно эту «регуляцию». Воспринято не было, вернее, было воспринято с возмущением – как это можно путать «регуляцию» с каким-то поганым серотонином, который еще неизвестно существует ли! Следующий удар нанесла Анна Самойловна, известная под ником «Танк». Она объяснила очень недовольным голосом, что, во-первых, то, что у нас получилось – неправильно, потому что не соответствует данным Дриша (чего, надо понимать, у нормального биолога случиться не может). Во-вторых, все полученное нами представляет собой повторение классических данных, а потому ничего нового и интересного в себе не содержит. Я попробовал было вякнуть, что давайте что-нибудь одно: либо неправильно, либо неново… Это тоже не возымело. Потом плохо помню, но вроде Васецкий более или менее дружелюбно объяснял мне мысли Анны Самойловны, а Нейфах иронизировал насчет слова «длинник», которым я пользовался для определения положения веретена, и спрашивал: есть ли слово «коротник», а я позорно забыл, что есть слово «поперечник». Общее ощущение после семинара было, как будто я вылез из-под асфальтового катка, и что я оказался совершенно неспособным объяснить и свои данные, и свою концепцию, а я-то думал, что красноречив… Слово «регуляция» возненавидел на всю оставшуюся жизнь. ГенСеич меня потом слегка пытался успокоить, сказав, что люди вообще плохо верят в медиаторы в эмбриогенезе, но чувствовалось, что он не слишком доволен ни моим дебютом, ни шумом вокруг семинара. Забавно, что позже Маша Скоблина сама стала заниматься ролью циклических нуклеотидов и кальция в созревании, что-то расспрашивала меня по этим вопросам, но никогда не возвращалась к тому семинару. Впрочем, не исключено, что никто, кроме меня, о нем и не помнит. Похоже, что потом к нашим данным просто привыкли, и они тоже стали чем-то само собой разумеющимся. Об этом можно судить по реплике, которую бросил Белоусов[2] на моей докторской защите: «Да, вот рассматривая эту работу, мы и вспоминаем, кто это, собственно, сделал…» * * * Семинар ясно продемонстрировал, что мы поохотились в чужих угодьях, да еще и явно оскорбили священное писание, повредили тотемы и осквернили капища эмбриологов. Надо было с этим разбираться всерьез, раз я хотел отквитаться за учиненный мне погром, тем более, что я собирался заниматься этой темой и дальше. Полез в библиотеку, нашел первоисточник. Классик, действительно выдающийся человек – Ханс Дриш в 1891 году опубликовал статью длиной в 119 страниц в Zeitschrift f?r Wissenschaftliche Zoologie. Беда в том, что статья была написана на немецком, который я не только не знаю, но и с детства не люблю. Вероятно, это наследственное, потому что моему папе пятерку в отличный аттестат поставили только с условием, что он больше никогда немецким заниматься не будет. Тем не менее, пробиваясь через немецкие словеса по полстрочки длиной, я кое-как смысл затеи, в той мере, в какой это меня касалось, вроде бы понял. Что не понял – объяснил ГА, который был в школе настоящим отличником, в том числе по немецкому, и даже на английском, который учил уже в Универе, говорил c явственными следами этого. В самом общем виде классика в этой области – это опыты Ру и Дриша. Вильгельм Ру опубликовал свои опыты в 1885 году – он горячей иглой убивал один из бластомеров лягушки на 2-х клеточной стадии, и убитую клетку не удалял, а потом из такого зародыша формировалась «половина» личинки. У Дриша получилось по иному – он отделял 2 из четырех клеток зародыша морского ежа, и у него образовывался полноценный зародыш. После того, как МакКлаллан изолировал блатомер лягушки и подтвердил данные Дриша, опыт Ру списали в артефакты, каковым он и числится в самом современном учебнике по биологии развития (Гёрдона). Это – «в самом общем», а в подробностях выяснились интересные дела! Во-первых, у Дриша при изоляции сначала получалась полубластула (полусфера, открытая своей полостью во внешнюю среду), которая только потом замыкалась. То есть, получается, что половинный зародыш у него сначала развивался именно как половина целого, и никаких последствий изоляции здесь вообще нет! Это зародыш потом сам, неизвестно от какой причины, восстанавливал сферическую замкнутую форму (что и именуется «регуляцией»). И с точки зрения ранних стадий, его результат нисколько от результата Вильгельма Ру не отличался! При этом опыт Дриша – это классика, а опыт Ру – артефакт! Я эти полубластулы в своих опытах тоже наблюдал (странная штука – «полцыпленка», да еще плавает!), как и возможность их «регуляции» - тоже, но у меня в части случаев (особенно, когда микромеры формировались с задержкой) половинная бластула сразу формировалась как полноценная бластула половинного размера! С микромерами у классика – еще кучерявее. Согласно работе Дриша, растиражированной во множестве вариантов его последователями, половинные зародыши всегда формируют два микромера! Однако, в той же самой статье рядом с рисунком половинного зародыша, содержащего два микромера, есть еще один рисуночек – на нем 16 клеток с четырьмя микромерами, но размер вроде бы половинный. И в тексте объяснение, что это, вроде, микрозародыш, хотя никаких доказательств того не приведено. Мужик-то он был добросовестный и факты описывал честно, но на мелочах не заморачивался – игнорировал, поскольку для него, видимо, в тот момент важен был сам факт способности изолированного бластомера сформировать полноценный зародыш. Откуда взялись наши с Дришем расхождения мне удалось разобраться существенно позже. И я очень хорошо помню, что начал эту работу спустя 86 лет после него и потратил на то, чтобы разобраться 30 с лишним лет. В любом случае и Ру, и Дриш были в мировой науке первыми, кто додумался до гениально простых опытов, и создали основу биологии развития. Анализ результатов и выводы из вроде бы очевидных результатов могут и меняться с получением новых знаний. А вот то, во что превратили наследие Дриша – это классика некритического восприятия, преклонения перед авторитетом и нетерпимость к новым фактам и тем, кто их по невежеству своему получил. Еще в конце 20-х известный эмбриолог Плаф (Plough) попытался повторить опыты Дриша и констатировал, что, так, как у отца-основателя, у него не получилось – полноценные половинные зародыши формировались далеко не из всех изолированных бластомеров. Характерно, какой вывод из этого дискрипанса сделал сам Плаф: - Вероятно, моя техника изоляции бластомеров уступает технике Ханса Дриша. Ну, во-первых, это – странноватая реклама собственных кривых рук, а, во-вторых, отсутствие даже попытки понять, в чем же, собственно, состоят отличия… Только в конце 70-х американская исследовательница, видимо, уже не страдающая преклонением перед авторитетами, повторила опыт Дриша и прямо написала, что только около 20% половинных зародышей формируют квазинормальную личинку…
Что касается раннего развития, то на основе данных Дриша, по меньшей мере с конца 30-х гг., возникла целая концепция «микромерных часов» еще одного классика эмбриологии – Гёрстадиуса. Суть ее сводится к тому, что при любых экспериментальных воздействиях (разрезание одноклеточного зародыша по любой оси, кроме экваториальной, изоляция бластомеров) микромеры формируются точно в тот срок, когда это происходит у интактных зародышей. Картинка из статьи Гёрстадиуса 1939 года так и кочует по сию пору из статьи в статью, из учебника в учебник (один из них был даже прислан мне в подарок последователем Гёрстадиуса – Жигаком). И ведь и здесь были люди, которые примечали, что не все так гладко в классических данных – ведь американская специалистка по морскому ежу Харвей в 40-м году писала, что количество микромеров у половинных зародышей может быть разным, а потом, видно, авторитетом ее задавили, и в ее компендиуме по морскому ежу Arbacia punctulata в 56-м году уже написано, что «у половинных зародышей ВСЕГДА по 2 микромеры» - в тексте это особо выделено… Так что мы со своими коррективами, перпендикулярными классике, пришлись не ко двору и нарушили сложившееся благолепие. За это я и поплатился на том семинаре… Только летом 2007-го года я отыгрался по-полной – на симпозиуме по морфогенезу сделал доклад при полном зале эмбриологов, проанализировал классические и собственные данные, последних уже было предостаточно, проехался по их предубеждениям и священным коровам, и никто уже не пытался меня закатать в дорожное покрытие. Потому что крыть было нечем… Все просто… На следующий год Чайлахян со товарищи отправились на Витязь немного раньше меня. Даром времени они не теряли – Левон выяснил, что на плоском морском еже бластомеры без всяких усилий можно разделять тонкой стеклянной иглой, которую вытягивали на спиртовке из капилляров, которые используются для приготовления микроэлектродов. При этом, чтобы научиться делать иглы, пришлось потратить больше времени, чем на то, чтобы научиться резать зародыши. Во всяком случае, уже за второй день я нарезал больше бластомеров, чем за весь прошлый сезон. Потом результаты росли, за один опыт я иногда успевал получить (до и после адгезии) свыше 120 бластомеров, и это давало возможность ставить вполне массовые серии с фармакологией и обязательными контролями. В тот год (1978-й), как будто специально, материал был очень высокого качества, а типы дробления раскладывались просто классически – большинство изолированных до адгезии бластомеров микромеров на 4-м делении дробления не формировало, а после – дисциплинированно формировали. Вопрос о том, кто же все-таки прав здесь – мы или Дриш – решился сам собой. Пока экспериментальных бластомеров была сотня – еще можно было сомневаться, а когда более 4000 – уже нельзя. На этом материале мы с Володей Смоляниновым нарисовали красивую схемку: клетка на каждом делении получает или не получает сигнал о наличии соседки, а потом вступает в действие фактор отличия свойств цитоплазмы на вегетативном полюсе, и при достижении некоторого ее критического уровня формируется асимметричное веретено и, соответственно, выделяются микромеры. Эта идея и посейчас актуальна. Причин наших с Дришем расхождений, скорее всего, две. Во-первых, это банальное различие видов, на которых мы ставили опыты. В 89-м в Которе я дорвался до того самого Paracentrotus lividus, на котором работал Дриш, и тут меня ждали сюрпризы. Во-первых, вопреки майсам, которые распространял Бовери, никакого пурпурного пояска на вегетативном полюсе там нет – только какие-то хаотически разбросанные пятнышки… А, главное, и в Которе, и много позже в Неаполе оказалось, что у множества партий зародышей микромеры могут формироваться уже на 8-ми клеточной стадии, а не на 16-ти клеточной. Еще бы у Дриша в такой ситуации не наблюдались микромеры на 4-м делении дробления! А вот если смотреть микромеры у половинных зародышей на 3-м – так там картина полностью повторяет то, что творится на плоском и черном ежах! Кстати, на плоском еже преждевременные микромеры тоже не редкость – надо постоянно присматривать за 8-клеточной стадией. Тогда в Черногории на Paracentrotus успех, надо признать, получился половинчатым, потому что изолировать бластомеры до адгезии не очень получалось – в нормальной воде это было невозможно из-за жесткого гиалинового слоя, а в бескальциевой большинство изолированных бластомеров сразу же разрушалось. Только в 2006-м в Неаполе я допер до простой вещи – как и для измерения мембранных токов надо переносить зародыши из бескальциевой воды в нормальную непосредственно перед изоляцией. Помогло! Гиалиновый слой в достаточной мере восстановиться не успевал, а кальциевого шока произойти не могло, и распад типов дробления сразу получился. Что приятно. Вторая причина расхождений с классиком – поглубже и заключается в самом механизме взаимодействия. Можно думать, что Дриш как настоящий немец был отменно аккуратен и опыты ставил, строго соблюдая процедуру – то есть, изолировал бластомеры всегда в одно и то же время – после того, как процесс клеточного деления был полностью завершен. Естественно, что при таком подходе и результат у него получался строго униформный. Мы же поначалу изолировали, когда бог на душу положит, потому что, во-первых, разгильдяи и не думали о соблюдении строго стандартных условий опыта, а, во-вторых, При таком количестве материала и практически готовой экспериментальной модели, которую мы стали называть «микромерной», можно было развернуть и опыты, которые меня, собственно, больше всего и интересовали – исследовать возможность трансмиттерного контроля межклеточных взаимодействий. Запахло возможностью опровергнуть постулат Бузникова о том, что трансмиттеры сами по себе не могут быть межклеточными посредниками в эмбриональных взаимодействиях. Cеротонин исправно увеличивал число половинных зародышей с микромерами, а его антагонисты – давали противоположный эффект. Удалось даже провести серию на специфичность эффекта – с добавлением одновременно серотонина и антагонистов с соответствующими контролями. Впереди открывался путь экстенсивного развития – набора кучи фармакологических данных. Однако, как и почти всегда в это время, пришел тайфун, качественный материал исчез, а КПД опытов упал раза в три. Ничего не поделаешь, пришлось смириться и ждать следующего сезона. (Продолжение следует)
|

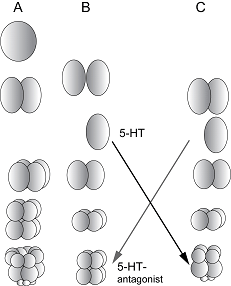 потому что старались нарезать как можно больше бластомеров, начиная с момента завершения борозды дробления. Редкий случай, когда поспешность плодотворна, она-то и принесла «неклассический» результат.
потому что старались нарезать как можно больше бластомеров, начиная с момента завершения борозды дробления. Редкий случай, когда поспешность плодотворна, она-то и принесла «неклассический» результат.