| Гостевая | yurishmukler@yahoo.com | Ю. Б. Шмуклер | Галерея | О себе | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Новости | Летопись текстов | Из дальних странствий | Моя семья и другие звери | |||||||||
| Биологическое | ЦСКА | Друзья и родственники | Генеалогия | Бреды и анекдоты | ||||||||
|
||||||||||||
|
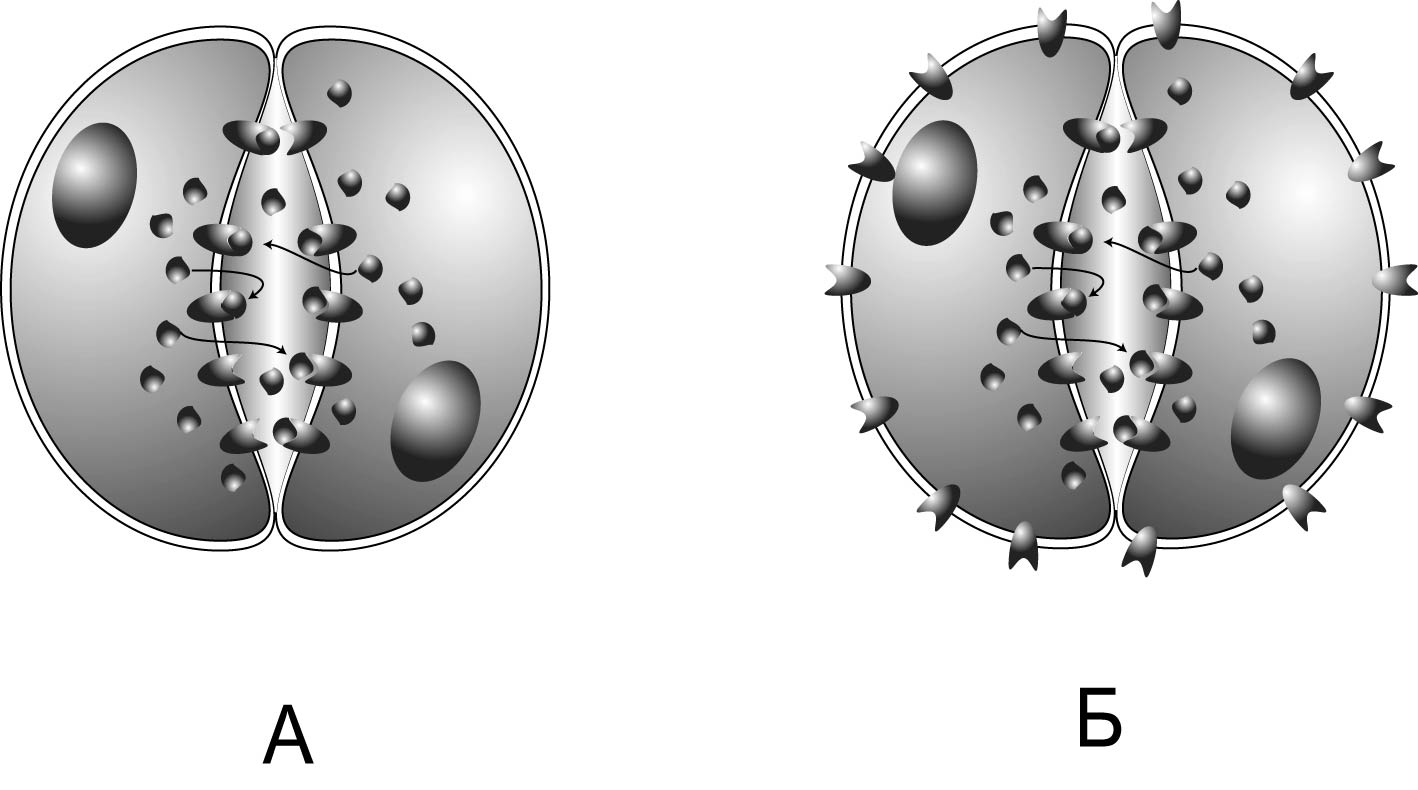
Протосинапс Модель, которую я придумал в конце 80-хх годов, и впервые опубликовал на русском в журнале «Биологические мембраны» (Шмуклер, 1992), а потом и на английском (Shmukler, 1993), описывает возможный механизм взаимодействия бластомеров в период делений дробления (и может представлять собой, с одной стороны, отражение первичного механизма межклеточных взаимодействий в эволюции, а с другой – субстрат развития собственно синаптических механизмов) Модель основана на том, что концентрация серотонина в межбластомерной щели повышена (Markova et al., 1985) вследствие выброса медиатора из бластомеров во внешнюю среду (таков механизм его инактивации, поскольку моноаминоксидазы у ранних зародышей слабы) и того, что сестринский бластомер и изолирующие межбластомерную щель контакты представляют собой пассивное препятствие для его утечки во внешнюю среду. Таким образом, вероятность взаимодействия медиатора с соответствующими рецепторами в контактной области выше, чем на свободной поверхности бластомеров, где диффузия медиатора не встречает препятствия. Присутствие серотониновых рецепторов (вероятно, 3-го типа) на поверхностной мембране бластомеров морского ежа подтверждается данными связывания меченных лигандов, электрофизологическими, эмбриофизиологическими данными, а также результатами исследований с помощью флуоресцентных зондов (Shmukler, 1993; Shmukler et al., 1999; Shmukler , Tosti, 2002). Таким образом, предполагается существование «протосинапса», в котором оба бластомера в ходе первого деления дробления являются как источником, так и мишенью медиатора, т.е. совмещают в себе функции пре- и постсинаптической клетки.
Аллегория протосинапса
|